

Vyšlo nové číslo Botaniky – 1/2026
V jarním čísle se dočtete např. o rostlině roku 2026 hořci brvitém, o slanomilných houbách či o jedné mimořádně zajímavé strategii rostlin. Díky fotoreportáži získáte představu, jak vypadá práce českých botaniků v Jižní Africe, a nechybí ani rozhovor – tentokrát...
Publikováno: 20 dubna, 2026
více...
Pražské oázy biodiverzity
Mohlo by se zdát, že v hustě osídleném území metropole už přírodovědce nic nepřekvapí. Přesto se ukazuje, že i v Praze se stále dají objevit druhy, které dosud nebyly z České republiky známy. Nedávné průzkumy vědců Botanického ústavu AV ČR...
Publikováno: 20 dubna, 2026
více...
Známý neznámý hořec brvitý aneb rostlina roku 2026
Rostlinu roku vyhlašuje Česká botanická společnost letos již pošesté. Po invazním starčku úzkolistém, který byl rostlinou roku 2025, titul pomyslně přebírá hořec brvitý, původní, avšak ustupující druh naší květeny. Volbou tohoto druhu se ČBS připojuje k Roku hořců, letošní popularizační...
Publikováno: 20 dubna, 2026
více...
Strategie, která není pro každého
Představme si louku po seči. Tráva je skoro pryč, byliny jsou useknuté, z barevné louky zůstaly jen zbytky stébel a stonků a trocha žlutozeleného listí. Z dálky to vypadá, jako by se většině rostlin jejich život náhle hodně zkomplikoval. A...
Publikováno: 20 dubna, 2026
více...
Sůl nad zlato aneb tajuplný život hub v osmoticky náročném prostředí
Představme si louku po seči. Tráva je skoro pryč, byliny jsou useknuté, z barevné louky zůstaly jen zbytky stébel a stonků a trocha žlutozeleného listí. Z dálky to vypadá, jako by se většině rostlin jejich život náhle hodně zkomplikoval. A...
Publikováno: 20 dubna, 2026
více...
Život na hraně: biologické půdní krusty v Arktidě
Když se řekne poušť nebo arktická tundra, většina z nás si představí krajinu, kde příroda zmáčkla tlačítko „pauza“: žádné stromy, jen pár keříků, písek, kameny a vítr. Místa, která vypadají, jako by to tady život skoro vzdal. Jenže skutečnost je...
Publikováno: 20 dubna, 2026
více...
Za rostlinami Jižní Afriky
Nejjižnější cíp Afriky patří k botanicky nejzajímavějším místům na světě. Na relativně malé ploše roste neobyčejné množství druhů, z nichž mnohé nenajdeme nikde jinde. Oblast je proto považována za jedno z hlavních světových center biodiverzity, a dokonce za samostatnou floristickou...
Publikováno: 20 dubna, 2026
více...
Třicet let ekologie obnovy u nás
V minulém roce uplynulo 30 let od vydání článku, který poprvé představil českým čtenářům tehdy se formující obor ekologie obnovy. Jaké poznatky výzkum přinesl a jaké jsou jeho dopady do praktické ochrany přírody a krajiny, přibližuje autor zmíněného článku Karel...
Publikováno: 20 dubna, 2026
více...
Pryskyřice jehličnanů a jejich výzkum v Průhonickém parku
Při studiu původu třetihorních jantarů ze severozápadních Čech a východní Moravy si autoři položili otázku, zda by klíčem k identifikaci jantarů, tedy fosilních pryskyřic, nemohlo být složení pryskyřic současných jehličnanů. Odpověď se rozhodli hledat v Průhonickém parku, který splňuje všechny...
Publikováno: 20 dubna, 2026
více...
Vyšlo nové číslo Botaniky – 2/2025
V podzimním čísle pokračuje série článků o novinkách v české flóře, dočtete se také o přezimujících pupenech, výmladkových lesích, štěrkopískových náplavech na Labi či o pátrání po cizích genech v genomech ječmenů. Detektivní pátrání připomíná i studium evoluční historie horských...
Publikováno: 7 listopadu, 2025
více...
Evropské ekosystémy reagují na oteplování rozdílně
Rostlinná společenstva v Evropě procházejí dlouhodobými proměnami. Druhy, které dávají přednost chladnějším podmínkám, postupně ustupují, zatímco teplomilnější druhy se více rozšiřují. Mezinárodní tým vědců s účastí odborníků z Botanického ústavu AV ČR nyní poprvé porovnal, jak tyto změny probíhají v...
Publikováno: 7 listopadu, 2025
více...
Co nového v české flóře 3
Článek přináší zprávy o nálezech nových druhů či kříženců, o nově nalezených zavlečených či zplanělých druzích, o nových lokalitách, kde byly dané druhy spatřeny poprvé či po mnoha letech. Článek v pdf ke stažení (427 kB) Detail klásku miličky zelenavá (Eragrostis...
Publikováno: 7 listopadu, 2025
více...
Horizontálně, prosím!
Když se řekne „dědičnost“, většina lidí si představí předávání dědičné informace z rodičů na potomky. Takové dědičnosti říkáme vertikální. V přírodě ale existuje ještě jiná, méně známá cesta – horizontální přenos genů. To je způsob, kdy si organismy předávají části...
Publikováno: 7 listopadu, 2025
více...
Mladí potomci, nebo dávní migranti?
Slovo jestřábník (latinsky Hieracium, čeleď Asteraceae) vzbuzuje u botaniků i dalších zájemců o přírodu obavy. Obvykle se při jeho vyslovení vybaví obrovská diverzita, množství obtížně rozpoznatelných druhů, polyploidie (přítomnost tří a více sad chromozomů) a apomixie, tj. vznik semen bez...
Publikováno: 7 listopadu, 2025
více...
Od pupenu k rostlině. Co nám prozradí přezimující pupeny?
Vytrvalé byliny tvoří podstatnou část naší středoevropské flóry. S příchodem zimy a prvních mrazíků sice nadzemní části bylin mizí z krajiny, ale život samotných bylin zdaleka nekončí. Naopak, pokračuje skrytě pod povrchem v půdě. Vytrvalost bylin znamená, že rostlina přežívá...
Publikováno: 7 listopadu, 2025
více...
Výmladkové lesy jako společné dílo přírody a člověka
Myslíme si často, že člověk přírodu jen ničí. Ve skutečnosti ale i mnoho cenných přírodních stanovišť vzniklo dlouhodobým působením člověka. Většinou se nám vybaví třeba květnaté louky, ale totéž platí i pro lesy. Výmladkové (pařezinové) hospodaření je krásným příkladem toho,...
Publikováno: 7 listopadu, 2025
více...
Postřehy ze života štěrkopískových náplavů
Úsek Labe mezi Ústím nad Labem a Hřenskem patří mezi přírodovědecky i krajinářsky nejzajímavější místa v České republice. Řeka si proráží cestu mohutnými kvádrovými pískovci a níže uloženým krystalinikem a tvoří až 300 m hluboký kaňon. Je to největší kaňon...
Publikováno: 7 listopadu, 2025
více...
Tradice českého ovocnictví a Pomologické arboretum na Chotobuzi
Průhonické pomologické arboretum, které se nachází v části obce zvané Chotobuz, navazuje na naše nejstarší ovocnářské tradice a je v současnosti nejstarším sbírkovým ovocnářským arboretem u nás. Jabloně v Pomologickém arboretu na Chotobuzi. Foto P. Sekerka
Publikováno: 7 listopadu, 2025
více...
Vyšlo nové číslo Botaniky – 1/2025
V letošním prvním čísle představujeme starček úzkolistý – rostlinu roku 2025, orchideje s nápadnými trojúhelníkovitými květy či škornice – bylinné příbuzné dřišťálů. Dále se dočtete o jednobuněčných zelených řasách krásivkách v Krušných horách nebo o strategii managementu invazních a nepůvodních...
Publikováno: 22 dubna, 2025
více...
Starček úzkolistý rostlinou roku 2025
Česká botanická společnost vyhlašuje každoročně rostlinu roku. Pro rok 2025 byl vybrán starček úzkolistý, vědeckým jménem Senecio inaequidens. Nejeden příznivec české květeny byl po oznámení letošní rostliny roku možná překvapen, nebo dokonce zklamán. Namlsali jsme se v minulých letech vzácných...
Publikováno: 22 dubna, 2025
více...
Rod Masdevallia a jemu příbuzné – orchideje s nápadnými trojúhelníkovitými květy
I když se orchideje vyskytují skoro všude na zemi, většina lidí je má spojené s tropickými pralesy. Není divu, v tropických oblastech je množství druhů orchidejí zdaleka nejvyšší a směrem k pólům klesá. Nejvíce druhů najdeme v tropech Ameriky. Roste...
Publikováno: 22 dubna, 2025
více...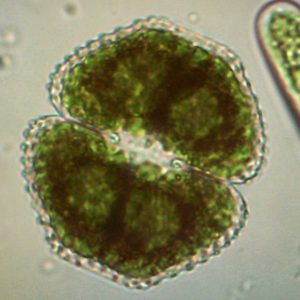
Krásivky krušnohorských rašelinišť v době klimatických extrémů
Krásivky čítají téměř 3300 druhů, přičemž z území České republiky je jich dosud známo více než 500. Díky své neobyčejné morfologické rozmanitosti jsou tyto druhy poměrně dobře detekovatelné rutinní mikroskopickou analýzou bentických vzorků. Nejen z tohoto důvodu se používají k...
Publikováno: 22 dubna, 2025
více...
Světlo a sucho v lese: Jak prostředí formuje pestrost bylinného patra?
Lesy přirozeně považujeme za jeden z klíčových nositelů biodiverzity. Mnoho druhů divoce žijících rostlin a živočichů je vázáno výhradně na lesy, nemluvě o obrovském bohatství lesních hub, mikroorganismů a dalších projevů života pod korunami stromů. Lesní stín však představuje stres,...
Publikováno: 22 dubna, 2025
více...
Půda pod lupou: jak rostliny ovlivňují své okolí
Půda je živý svět, který se neustále mění pod vlivem rostlin, mikroorganismů a dalších organismů. Zatímco řada studií se zabývá vlivem rostlin na půdu v jejich bezprostřední blízkosti, o vlivu rostlin na jejich vzdálenější okolí se ví stále poměrně málo....
Publikováno: 22 dubna, 2025
více...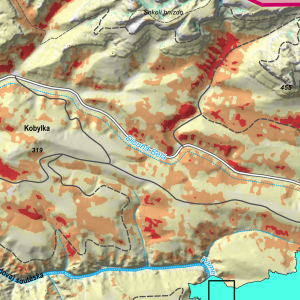
Mapy environmentálních proměnných: tolik potřebné, ale dlouho nedostupné
Podmínky prostředí – například teplota a vlhkost vzduchu či půdy, nadmořská výška, orientace a sklon svahu – zásadně ovlivňují rozšíření a rozmanitost organismů. Přestože mnohé české instituce již dlouhá léta sbírají a zpracovávají environmentální data, byla pro výzkumníky i širokou...
Publikováno: 22 dubna, 2025
více...
Strategie managementu invazních a nepůvodních druhů v NP České Švýcarsko a CHKO Labské pískovce
Přestože analýzy ukazují, že chráněná území fungují do určité míry jako bariéry proti šíření invazních druhů, ani jim se rostlinné invaze nevyhýbají. Příkladem je Národní park České Švýcarsko (NPČŠ) a k němu přilehlá Chráněná krajinná oblast Labské pískovce (CHKOLP). Jak...
Publikováno: 22 dubna, 2025
více...
Škornice – bylinné příbuzné dřišťálů
Dřišťálovité rostliny jsou známé především díky dřišťálům, keřům rozšířeným v mnoha druzích na severní i jižní polokouli. Málo se ví, že existují i bylinní zástupci této čeledi. Přitom bylinných rodů je více a jsou mnohem různorodější. Jednotlivé skupiny bylinných dřišťálovitých...
Publikováno: 22 dubna, 2025
více...
Za vodními a mokřadními rostlinami s novým atlasem
Článek představuje nový atlas vodních a mokřadních rostlin, který vznikl z iniciativy nestora české mokřadní ekologie Jana Květa jako dílo pracovníků Botanického ústavu a který připravuje k vydání Nakladatelství Academia.
Publikováno: 22 dubna, 2025
více...
Vyšlo podzimní číslo Botaniky – 2/2024
V novém čísle se dočtete o novinkách v české flóře, invazi kosmatců i o tom, jaký význam má neopadané listí. Dovíte se, jak poznat biologicky cenný les či jak rostliny v Himálajích reagují na změnu klimatu. Číslo uzavírá článek o...
Publikováno: 24 října, 2024
více...
Vyšel poslední díl Květeny ČR
Víte, co mají společného bér, bojínek, ježatka, kukuřice, metlice, pšenice, rosička, sveřep, válečka, vousatka, zaječí ocásek? Jsou to zástupci trav, v botanickém názvosloví čeledi lipnicovitých, ekonomicky a hospodářsky jedné z nejdůležitějších čeledí světa. Té je věnována největší část posledního, 9....
Publikováno: 24 října, 2024
více...
Co nového v české flóře 2
V této rubrice máte možnost seznámit se s významnými floristickými nálezy, které jsou publikovány v odborném časopise Zprávy České botanické společnosti (viz též Botanika 2/2023). Autoři edice Additamenta v nich referují o nálezech nových druhů či kříženců, o nově nalezených...
Publikováno: 24 října, 2024
více...
Invaze kosmatce: zrádná krása na pobřeží
Kosmatce mají atraktivní květy a rychle se klonálně šíří. Díky těmto vlastnostem byly introdukovány do mnoha oblastí po celém světě jako okrasné rostliny nebo ke stabilizaci půdy, zejména písečných dun. Od té doby osídlily miliony hektarů pobřežních oblastí po celém...
Publikováno: 24 října, 2024
více...
Nový vhled do evoluce a ekologie díky unikátní sbírce rostlin
Botanici se snaží lépe porozumět rostlinnému světu okolo sebe, proto tráví hodně času pozorováním rostlin v přírodě. Pozorování v přírodě však může být velmi náročné, protože druhy, které chceme zkoumat, mohou růst daleko od sebe, na nepřístupných exotických místech, jen...
Publikováno: 24 října, 2024
více...
Jak poznat biologicky cenný les?
Tuhle otázku si klade nejeden ochránce přírody, lesník a biolog. Společnost k tomu využívá různé způsoby. Význam lesa lze hodnotit např. podle stáří, množství mrtvého dřeva či jeho struktury. Použít ale můžeme i biodiverzitu, kterou zpravidla „měříme“, nebo spíše odhadujeme,...
Publikováno: 24 října, 2024
více...
Marcescence – přehlížený fenomén neopadaného listí
Každý podzim listí zezlátne a opadá, nebo alespoň jeho větší část. Ta menší zůstane na stromech nebo na suchých stoncích bylin. A když se na tento fenomén soustředíme, zjistíme, že neopadaného listí vlastně není zas tak málo. Přesto bylo vědci...
Publikováno: 24 října, 2024
více...
Jak půdní vlhkost chladí les
Teplota vzduchu během letního vedra je v podrostu lesa často o poznání nižší než mimo les. Les tedy má jistý chladící účinek, který není způsoben jen nižším množstvím dopadajícího záření. Prostřednictvím výparu se na něm může výrazně podílet i množství...
Publikováno: 24 října, 2024
více...
Himálaje – rostliny a změna klimatu
V Himálajích dochází k bezprecedentnímu nárůstu letních teplot, což má za následek nejen tání ledovců, ale mimo jiné také vegetační změny. Náš tým provádí výzkum v Himálajích, zejména v suché oblasti na západě pohoří, už od roku 2008, a tak...
Publikováno: 24 října, 2024
více...
Podzimní cibuloviny v Průhonické botanické zahradě
Rostliny mají různé strategie, jak přežít nepříznivé období. Jednou z nich je tvorba cibulí a hlíz. Velké množství živin, které je v nich uschované, jim umožní rychlý počáteční růst v okamžiku, kdy nastanou příznivé podmínky. Lužanka žlutá (Sternbergia lutea) je...
Publikováno: 24 října, 2024
více...
Vyšlo jarní číslo časopisu Botanika – 1/2024
V novém čísle se dočtete o rostlině roku 2024, druhově nejbohatších loukách či využití DNA při určování lišejníků. Dále se dovíte, jaký problém představují zvýšené hladiny hormonů v odpadních vodách a k čemu lze využít znalostí o polárních řasách. A...
Publikováno: 19 dubna, 2024
více...
Globální mapy současných i budoucích příčin narušení lesa
Zatímco tropické lesy budou podle předpovědi v budoucnu více sužovat požáry a větrné bouře, v severských lesích naopak může oheň a vítr hrát menší roli než dnes, větší vliv však budou mít škůdci a choroby. To je jeden ze závěrů...
Publikováno: 19 dubna, 2024
více...
Kosatec sibiřský rostlinou roku 2024
Česká botanická společnost vyhlašuje každoročně rostlinu roku. Cílem je upozornit na ohrožené nebo jinak zajímavé druhy naší květeny a seznámit širokou veřejnost s jejich biologií, ekologií a případně i jejich ochranou. Pro rok 2024 byl vybrán kosatec sibiřský, vědeckým jménem...
Publikováno: 19 dubna, 2024
více...
Co pamatují druhově nejbohatší louky
Už nějakou dobu se ví, že louky v Bílých Karpatech patří k druhově nejbohatším travinobylinným ekosystémům na světě – zejména pokud se zaměříme na počet druhů cévnatých rostlin na nevelkých plochách o rozloze metrů až desítek metrů čtverečních. Třeba na...
Publikováno: 19 dubna, 2024
více...
Sasanko, proč máš tak velké oddenky?
V mírném klimatickém pásmu je většina bylin vytrvalá. Na rozdíl od stromů a keřů vytrvalým bylinám zpravidla před začátkem zimního období odumírají nadzemní stonky. Vytrvalé byliny tak musí pravidelně obnovovat svá nadzemní pletiva po mrazivé zimě, někdy ale i po...
Publikováno: 19 dubna, 2024
více...
Jak spolehlivě identifikovat epifytické lišejníky
Tradiční výzkum biodiverzity se neobejde bez správné identifikace druhů. Kromě rutinních metod určování pomocí morfologických, anatomických a biochemických znaků se dnes běžně používají i metody založené na analýze DNA. Pro spolehlivou identifikaci druhů jsou však nutné referenční databáze takzvaných barkódových...
Publikováno: 19 dubna, 2024
více...
Genomová dominance aneb co se děje v genomech kříženců
Mezidruhová hybridizace (křížení) je proces, při němž potomky plodí rodiče, kteří nepatří ke stejnému biologickému druhu, někdy dokonce ani rodu. Ve srovnání s živočichy je takováto hybridizace u rostlin mnohem častější a významně přispívá ke speciaci, tedy vzniku nových druhů....
Publikováno: 19 dubna, 2024
více...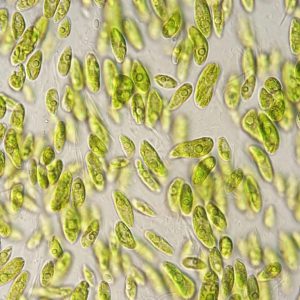
Polární řasy a jejich biotechnologický potenciál
Extrémní podmínky prostředí v polárních oblastech jsou výzvou pro všechny organismy. Primární producenti, kteří z anorganických látek produkují prostřednictvím fotosyntézy látky organické a tvoří základ potravních sítí, musí v polárních ekosystémech čelit mnoha stresovým faktorům. Patří mezi ně nízké teploty,...
Publikováno: 19 dubna, 2024
více...
Hormonální doping pro vodní ekosystémy
Spotřeba farmak celosvětově roste a s tím i jejich dopad na životní prostředí. Většina farmak totiž není odstraněna běžným technologickým procesem čištění odpadních vod a odtéká do vodních ekosystémů. Koktejl léčiv a dalších látek používaných naší civilizací, který analyzujeme na...
Publikováno: 19 dubna, 2024
více...
Z historie pěstování dřevitých pivoněk
Začátkem května začínají v Průhonické botanické zahradě kvést dřevité pivoňky. Jaký mají původ a co víme o historii jejich pěstování? Paeonia rockii ‚Su sha chan yi‘ – odrůdy pivoňky Rockovy se vyznačují výraznou tmavou skvrnou ve středu květu.
Publikováno: 19 dubna, 2024
více...
Novinky v Průhonickém parku
Tisíce jarních cibulovin, nové expozice v historických stavbách v parku, online mapa i pohádková trasa. V Průhonickém parku skončil projekt na jeho rozsáhlou obnovu, a návštěvníky tak čeká mnoho novinek. Rybárna v průhonickém parku. Foto M. Dvořáková
Publikováno: 19 dubna, 2024
více...
Vyšlo podzimní číslo časopisu Botanika – 2/2023
Nové číslo se věnuje novinkám v české flóře, schopnosti adaptace rostlin na nové klimatické podmínky, loukám či houbám. Přibližuje také, jak a proč vědci pátrají po původu nových druhů.
Publikováno: 25 října, 2023
více...
Skákající geny mají pozitivní vliv na rostlinná společenstva
Biodiverzita je spojována zejména s rozmanitostí druhů ve společenstvu, tzv. druhovou diverzitou. Vít Latzel z Botanického ústavu AV ČR se spolupracovníky se však při svém výzkumu zaměřili na rozmanitost uvnitř druhů, která může mít na fungování společenstva a ekosystému stejně...
Publikováno: 25 října, 2023
více...
Co nového v české flóře
V této nové rubrice máte možnost seznámit se s významnými floristickými nálezy, které jsou publikovány v odborném časopise Zprávy České botanické společnosti. Nové nálezy vychází ve Zprávách ČBS od roku 2002 pod názvem “Additamenta” (Additamenta ad floram Reipublicae Bohemicae). Jedná...
Publikováno: 25 října, 2023
více...
Schopnost adaptace rostlin na nové klimatické podmínky
Schopnosti rostlin rychle se geneticky přizpůsobit měnícím se podmínkám prostředí se říká rychlá evoluce. Je to fenomén, který v poslední době získává velkou pozornost, protože s rychle se měnícími podmínkami se setkáváme téměř na každém kroku. Tato schopnost je však...
Publikováno: 25 října, 2023
více...
Po stopách Charlese Darwina při hledání původu druhů
Charlesi Darwinovi trvalo více než 20 let od návratu z cesty kolem Jižní Ameriky, než výsledky svého výzkumu publikoval v knize „O původu druhů”. Uplynula další řada let a spousta výzkumné práce, než byly koncepty vytvořené Darwinem pochopeny. Dnes, téměř...
Publikováno: 25 října, 2023
více...
Čtyři desetiletí výzkumu krkonošských luk
Nedávno vyšla kniha o bádání na loukách v Krkonoších. Její základ vychází z dlouholetého výzkumu, který inicioval již koncem 70. let minulého století František Krahulec. Tři z autorů pracují v Botanickém ústavu v Průhonicích dodnes a stále se zkoumání luk...
Publikováno: 25 října, 2023
více...
Obnova druhově bohatých luk: proč mají některé druhy problém kolonizovat bývalou ornou půdu?
Středoevropské louky patří mezi druhově nejbohatší společenstva světa, alespoň pokud se díváme na plochy menší než přibližně 100 m2. Navíc hostí velké množství vzácných a zajímavých druhů rostlin, živočichů, hub a dalších méně známých organismů. Pokud však byly v minulosti...
Publikováno: 25 října, 2023
více...
Nelehký život mykorhizních hub v orných půdách
Pole zabírají významný podíl naší středoevropské krajiny a pro většinu organismů představují nepříznivé prostředí: jednotvárný porost, který na část roku mizí úplně, časté narušení agrotechnickými zásahy, aplikace chemických látek s cílem přímo eliminovat určité skupiny organismů. Tyto negativní vlivy mají...
Publikováno: 25 října, 2023
více...
Hniložijné houby jako brána mezi živými a mrtvými
Hniložijné (saprofytické) mikroskopické houby nalezneme na tlejícím dřevu, kůře, listech či stoncích na souši i ve vodě. Článek představuje zejména rod Chloridium, málo prozkoumanou a velmi rozmanitou skupinu hub, které sdílejí vzácný způsob tvorby výtrusů. Diverzita kolonií druhů rodu Chloridium....
Publikováno: 25 října, 2023
více...
Obnova Hlavní vyhlídky v Průhonickém parku
Botanický ústav AV ČR se kromě výzkumu věnuje také péči o Průhonický park. V posledních dvou letech tu Správa Průhonického parku provedla řadu úprav včetně obnovy hlavních kompozičních prvků. Článek přibližuje principy, podle kterých zakladatel Průhonického parku hraběte Silva-Tarouca tvořil...
Publikováno: 25 října, 2023
více...
Vyšla Botanika 1/2023
V prvním letošním čísle se dočtete např. o nově popsaném symbiotickém vztahu, o rostlině roku, tropických zázvorech či nepůvodních rostlinách v ČR. Nově bylo zařazeno několik rubrik a v čísle najdete rozhovor, fotoreportáž, pozvánky na akce i plakát.
Publikováno: 18 dubna, 2023
více...
Alkobioza – nově popsaný symbiotický vztah mezi houbami a řasami
Jan Vondrák z Botanického ústavu AV ČR se svými spolupracovníky popsal symbiotický vztah mezi houbami a řasami, který byl doposud ve vědeckém světě přehlížen. Pro soužití řas s kornatcovitými stopkovýtrusými houbami, které se zcela běžně vyskytuje v lesích mírného pásu,...
Publikováno: 18 dubna, 2023
více...
Kociánek dvoudomý rostlinou roku 2023
Česká botanická společnost vyhlašuje v pořadí již třetí rostlinu roku: po vstavači kukačce a hlaváčku letním se jí stává kociánek dvoudomý. Tato rostlina z čeledi hvězdnicovitých (Asteraceae), vědeckým jménem Atennaria dioica, je dalším příkladem druhu, který byl na území České...
Publikováno: 18 dubna, 2023
více...
Známé i neznámé tropické zázvory
Když se řekne zázvor, mnozí z nás si vybaví štiplavou, ale hřejivou chuť zázvorového čaje, ke kterému usedáme zejména při rýmě či nachlazení. Milovníci asijské kuchyně pak ocení chutě zázvoru, kurkumy, kardamomu nebo galangy v čínském, indickém nebo thajském pokrmu....
Publikováno: 18 dubna, 2023
více...
O nepůvodních rostlinách v Česku po dalších deseti letech
V prosinci 2022 vyšlo v časopise Preslia třetí vydání Katalogu nepůvodních rostlin České republiky, které navazuje na vydání z let 2002 a 2012. Bylo aktualizováno o nové údaje shromážděné v posledním desetiletí. Novinky v katalogu i v rozšíření nepůvodních rostlin...
Publikováno: 18 dubna, 2023
více...
Klíč k úspěchu invazních rostlin: půdní banka semen
Jednou ze základních otázek invazní biologie je, které z vlastností přispívají k úspěšnému zdomácňování nepůvodních druhů v novém prostředí (naturalizaci). Naturalizované druhy se totiž později mohou dále šířit a stávat se tak invazními. Výzkumy v poslední době ukázaly, že mezi...
Publikováno: 18 dubna, 2023
více...
Požáry a jejich vliv na vývoj lesa v minulosti i dnes
Národní park České Švýcarsko zažil v létě 2022 největší lesní požár, který kdy byl v naší moderní historii zaznamenán. Oheň pohltil celkem 1300 ha smrkových, borových a bukových lesů a bohužel zasáhl i několik domů. V důsledku dlouhodobého sucha a nahromadění snadno hořlavého...
Publikováno: 18 dubna, 2023
více...
Jak mikroorganismy rhizosféry ovlivňují rozšíření rostlin v Himálaji
Rozšíření rostlin je mimo jiné ovlivněno i jejich vztahy s půdním mikroorganismy. Ačkoli mají zásadní význam pro vývoj vegetace, jsou tyto vztahy stále málo prozkoumané, zejména ve vysokohorských ekosystémech. Rozhodli jsme se je studovat v Himálajích, které jsou ideální přírodní...
Publikováno: 18 dubna, 2023
více...
Kosatce – rostliny nejen krásné, ale i užitečné
Kosatec je nepochybně krásná květina zdobící by nepotěšila kytice těchto pestrobarevných květů s výraznou vůní? Za krásnou tváří se však skrývají nečekané účinky. V rámci naší studie jsme se zaměřili na vytipování perspektivních druhů a odrůd na základě testování bioaktivních...
Publikováno: 18 dubna, 2023
více...
Místo botanických zahrad v komplexní druhové ochraně rostlin
Botanické zahrady nejsou jen krásná místa pro příjemné strávení dne. Jsou to i důležitá místa, která předvádějí rozmanitost rostlin a umožňují výzkumníkům najít nové způsoby, jak chránit a zvyšovat biologickou rozmanitost. Podle aktuálně publikované studie v časopise Nature Plants botanické...
Publikováno: 18 dubna, 2023
více...
Alpinum Průhonického parku – Fotografický atlas
V novém speciálu časopisu Botanika se seznámíte se „starým“ Alpinem nad Podzámeckým rybníkem a „hlavním“ (nejstarším) Alpinem s významnými dřevinami a pamětihodnostmi. Každá část je členěna podle místních pojmenování převzatých po generací zahradníků a samozřejmě nechybí ani články o jeho ...
Publikováno: 6 února, 2023
více...
Vyšlo druhé letošní číslo Botaniky
Vážené čtenářky a vážení čtenáři, jsme tu s podzimním číslem Botaniky. Po přiblížení polistopadové historie našeho pracoviště zalétneme ještě na chvíli do exotiky jižní Afriky za rodem Pteronia. Z cest si člověk ale může přivézt i např. invazní druhy rostlin....
Publikováno: 13 listopadu, 2022
více...Porevoluční změny v historii Botanického ústavu
V minulém čísle jsme představili vznik Botanického ústavu a nyní poodhalíme změny, které nastaly po sametové revoluci. Soustředěné úsilí všech porevolučních ředitelů včetně současného Jana Wilda vedlo v přetvoření našeho pracoviště ve špičkovou vědeckou instituci spadající do nejlépe hodnocené kategorie...
Publikováno: 13 listopadu, 2022
více...
Za rodem Pteronia do Jižní Afriky a Namibie
Během našich cest do Jižní Afriky (především kapské květenné říše), která nás dlouhodobě fascinuje svým rostlinným bohatstvím, jsme narazili na rod, který jsme z našich končin neznali. Vzhledem k více než 9 tisícům druhů kapských cévnatých rostlin, z nichž 68...
Publikováno: 13 listopadu, 2022
více...
Dva odstíny šedi aneb Odhalování ekologie „nových“ vetřelců
Nedávné průzkumy zámeckých zahrad a městských parků ukázaly, že se ze zahradních výsadeb mimo obvyklé parkové dřeviny, jako je šeřík (Syringa vulgaris), ořešáky (Juglans nigra, J. regia), jírovec (Aesculus hippocastanum) či akát (Robinia pseudoacacia) šíří také množství druhů bylinných. V...
Publikováno: 13 listopadu, 2022
více...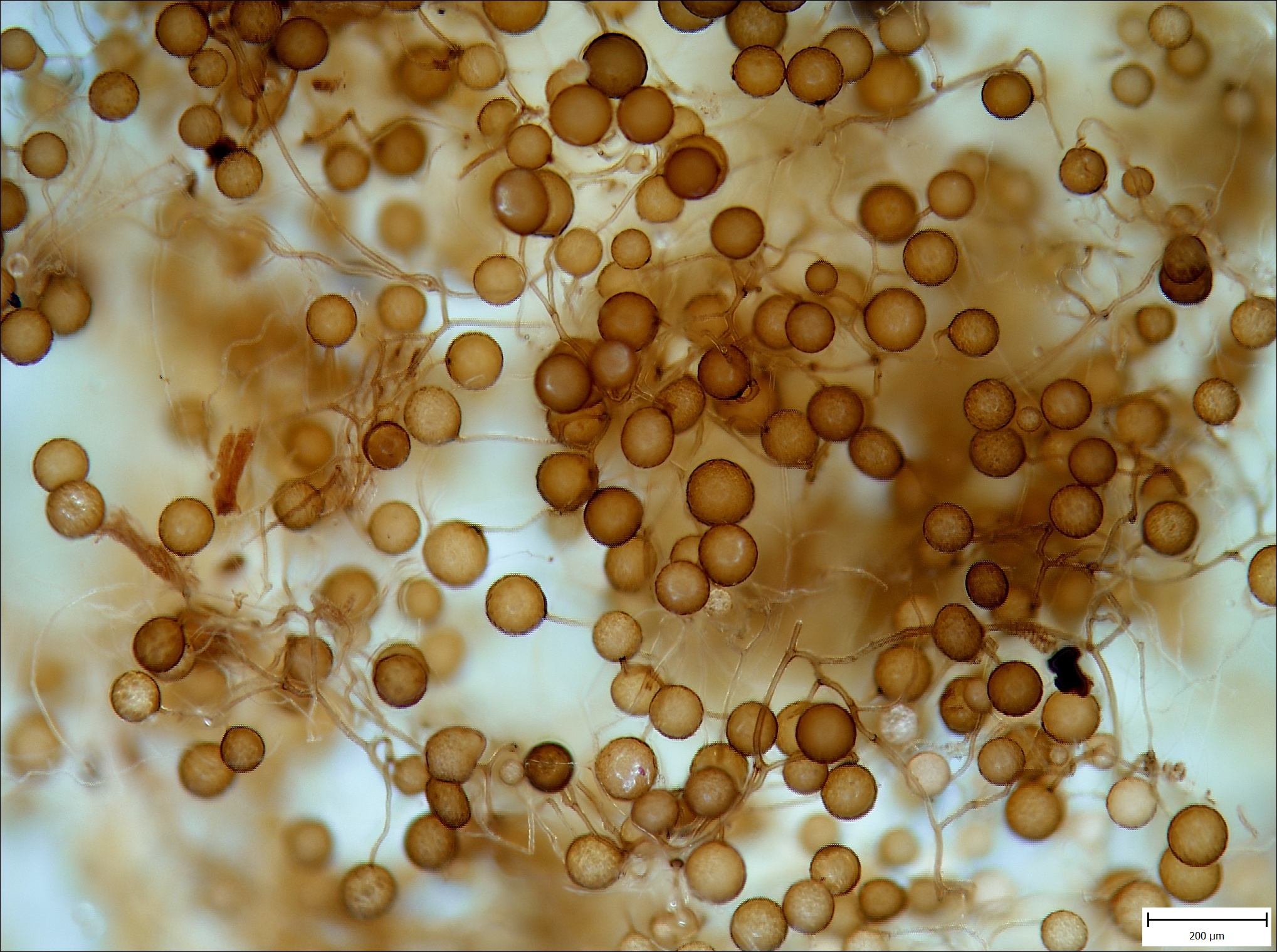
Má každá rostlina nejradši jen to svoje místečko?
Už po mnoho let je lidem známo, že není dobré rok co rok na jednom políčku pěstovat stále tu samou plodinu. Dokonce už před více než 2000 lety lidé zjistili, že ovocné stromy špatně rostou na místech, kde byly předtím...
Publikováno: 13 listopadu, 2022
více...
Na začátku byla sinice! Jak sinice (a řasy) formovaly život na naší planetě?
Sinice, skupina fotosyntetických bakterií, která v tuto chvíli čítá téměř 2 300 druhů ve zhruba 450 rodech, nás překvapí svou tvarovou různorodostí a ještě více svojí schopností osídlit prakticky všechna prostředí na Zemi, včetně těch extrémních, jako jsou termální prameny...
Publikováno: 13 listopadu, 2022
více...
Milířiště a lesy dnes a v minulosti
Dřevěné uhlí zná každý, kdo se už zúčastnil víkendového grilování na chalupě. Mnozí také vědí, že tento materiál hrál v minulosti velmi důležitou roli při různých industriálních aktivitách. Jednoduše řečeno, dřevěné uhlí má jednu vlastnost, která ho činila takřka nenahraditelným...
Publikováno: 13 listopadu, 2022
více...
Ohrožená diverzita lužních lesů
Ztráta biodiverzity představuje jeden z nejpalčivějších problémů naší planety. Týká se téměř všech ekosystémů a ušetřeny nezůstaly ani středoevropské lesy, lužní porosty nevyjímaje. V letech 2019–2021 jsme hodnotili změny diverzity vegetace nížinných lužních lesů České republiky za použití metody opakovaných...
Publikováno: 13 listopadu, 2022
více...
Novinky v Průhonické botanické zahradě
Letošní sezónu jsme zahájili dříve, než bylo v minulých letech obvyklé. Sezóna u nás začala s pomology z ČSOP jarním seminářem řezu ovocných dřevin. Dlouhodobě s nimi spolupracujeme na údržbě pomologického arboreta tak, aby arboretum bylo udržováno co nejšetrněji. Pomologické...
Publikováno: 13 listopadu, 2022
více...
Botanická zahrada Třeboň v roce 2022
Stálé a sezónní expozice naší botanické zahrady čítají už více než 1 000 druhů rostlin, většinou z mokřadních stanovišť různých koutů světa. V letošním roce jsme se zaměřovali na spolupráci s orgány ochrany přírody při záchraně některých kriticky ohrožených druhů...
Publikováno: 13 listopadu, 2022
více...
Vyšlo 1. letošní číslo Botaniky 1/2022
Vážené čtenářky a vážení čtenáři, je tu opět jaro a s ním kromě letošního 60. výročí našeho pracoviště vstupuje časopis Botanika do 10. roku své existence. To vybízí k malému ohlédnutí. Z celkem 233 příspěvků mezi roky 2013 a 2021...
Publikováno: 23 května, 2022
více...Jak šel čas v Botanickém ústavu do roku 1989
V lednu tohoto roku oslavil Botanický ústav AV ČR 60 let od svého založení. Za šest dekád své existence se z malých geobotanických pracovišť stalo největší centrum botanického výzkumu v ČR, které významně přispívá k výzkumu rostlin a ochraně přírody...
Publikováno: 23 května, 2022
více...
Co se lze dozvědět z dlouhodobých sledování aneb půl století se skalní stepí
Také si rádi listujete starými novinami, třeba i z doby vašeho dětství? S úžasem sledujete, kolik v dané době stál litr mléka a co bylo zásadním společenským tématem? I my botanici máme svoje „staré noviny“. Jsou jimi starší záznamy o...
Publikováno: 23 května, 2022
více...
Quo vadis, Adonis aestivalis?
Česká botanická společnost zvolila pro rok 2022 rostlinou roku hlaváček letní (Adonis aestivalis). Rod hlaváček (Adonis) patří do čeledi pryskyřníkovitých a zahrnuje přes 30 druhů (doma je v Evropě, Asii a Severní Africe, ale zavlečen byl i do Severní Ameriky...
Publikováno: 23 května, 2022
více...
Podobnost neznamená příbuznost a naopak: případ lakušníků
Lakušníky jsou vodní rostliny s podivuhodně spletitou evoluční historií. Jednotlivé druhy dokáží „klamat tělem“: jejich vzájemná podobnost může být v příkrém rozporu s evoluční vzdáleností mezi jednotlivými druhy. Více v článku: Prančl J. (2022): Podobnost neznamená příbuznost a naopak: případ...
Publikováno: 23 května, 2022
více...
Za kytkami s mobilem v ruce
Naši pradávní předkové se při hledání vlastní potravy museli mít neustále na pozoru, aby se sami nestali potravou. V dnešní krajině, zbavené velikých predátorů, máme pro sledování přírody většinou už jiné důvody. Chceme ji poznávat nebo v ní prostě jen...
Publikováno: 23 května, 2022
více...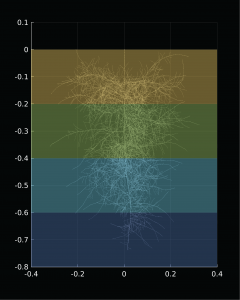
Rostliny jsou prostředníky mezi půdou a atmosférou
Mezi zemským povrchem a atmosférou dochází k nepřetržité výměně hmoty a energie, která přispívá k vývoji lokálního počasí i globálního klimatu. Svým působením tuto výměnu významně urychlují rostliny, které kořeny přijímají vodu z půdy, aby ji pak z nadzemních částí...
Publikováno: 23 května, 2022
více...
Život pro kuřičku – jak spolupráce mezi vědci, ochránci přírody a místními obyvateli zachránila českého endemita
Kuřička hadcová je na první pohled nenápadná rostlina z čeledi hvozdíkovitých, u které jako by nic nenapovídalo tomu, že patří mezi nejvzácnější české rostlinné druhy. S kuřičkou se již naši čtenáři mohli seznámit na stránkách Botaniky (2016/2). Kuřička je českým...
Publikováno: 23 května, 2022
více...
Cesta za poznáním a záchranou endemické mateřídoušky ostrova Mallorca
Kurátorky Průhonické botanické zahrady navštívily během své únorové týdenní stáže botanickou zahradu Jardí Botànic de Sóller v městečku Sóller ležícím na severozápadním pobřeží ostrova Mallorca ve Španělsku. Cílem jejich cesty bylo zdokonalení praktických dovedností, které později využijí při řešení aktuálních...
Publikováno: 23 května, 2022
více...
Ohlédnutí za Expo v Dubaji, kde čeští vědci přeměnili vodu ze vzduchu
Téměř dvě stovky pavilonů s účastí více než 190 zemí, které navštívilo 23 miliony návštěvníků. Tak by se dala shrnout světová výstava Expo 2020 v Dubaji, která skončila letos v březnu. Český pavilon přitáhl pozornost téměř 1,3 milionu návštěvníků. V...
Publikováno: 23 května, 2022
více...
Fotografický průvodce Průhonickým parkem – Listnaté stromy 2
Vyšlo speciální číslo časopisu Botanika zaměřené na listnaté stromy druhé části Průhonického parku, tzv. Obory. Kromě fotografií a přehledových map obsahuje i popisy 50 rodů dřevin s vyznačením více než 170 taxonů, které se v parku vyskytují. Průvodce můžete zakoupit...
Publikováno: 6 února, 2022
více...
Vyšlo 2. číslo Botaniky v roce 2021
Je tu podzim a s ním druhé číslo Botaniky. Léto bylo deštivé, a tak jsme mohli studovat polní mokřady, o kterých se dočtete v článku z míst, která v tomto rozmarném létě zasáhlo tornádo. Podobné atmosférické jevy o sobě dávají...
Publikováno: 15 listopadu, 2021
více...
Jednoho deštivého léta, aneb Co jsme (zatím) vybádali v polních mokřadech jižní Moravy
Bažiny, močály, mokřiny, třasoviska, mokřady… tato slova u části veřejnosti vzbuzují odpor či úzkost, patrně kvůli předpokládanému riziku utonutí nebo přinejmenším zamokření a zablácení.
Publikováno: 15 listopadu, 2021
více...
Velké sopečné erupce snižují aktivitu tropických cyklón
Velké sopečné erupce, jakými byly např. výbuch Tambory (1815) či Mount St. Helens (1980), ovlivňují globální klimatické podmínky. Velké množství aerosolu síranů vyvrženého do stratosféry, kde se dokáže udržet několik let, způsobuje citelné snížení teplot, které v minulosti zapříčinilo nízkou...
Publikováno: 15 listopadu, 2021
více...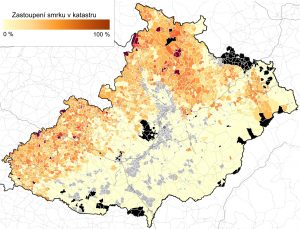
Jak zobrazit lesní minulost?
Na Oddělení vegetační ekologie Botanického ústavu AV ČR vědci v průběhu posledních deseti let nashromáždili velké množství informací týkajících se lesní minulosti. Jeden z dílčích výsledku je díky spolupráci s Ústavem pro hospodářskou úpravu lesa (ÚHÚL) volně k prohlížení v...
Publikováno: 15 listopadu, 2021
více...
Jak ve výzkumu propojovat archeologii a botaniku?
Archeologie je vědní disciplína, která se zabývá zkoumáním a uchováváním hmotných pozůstatků lidských společností z minulosti. Tyto lidské společnosti (stejně jako současné) však nelze poznávat odtržené od jejich životního prostředí, a proto se je archeologie snaží pochopit v jejich environmentálním...
Publikováno: 15 listopadu, 2021
více...
Mikroskopické šperky ze sbírky vodních a mokřadních rostlin v Třeboni
Botanické a stejně tak i zoologické zahrady jsou často refugiem nejen pro cílené druhy organismů, ale i pro další organismy přítomné, které je osídlily tak trochu „mimo plán“. Sbírka vodních a mokřadních rostlin v Třeboni hostí ve svých nádržích kromě...
Publikováno: 15 listopadu, 2021
více...
Botanická zahrada Třeboň 2021
Botanická zahrada Třeboň zahájila letos provoz tradičně 1. května a do konce září přilákala přes 2600 návštěvníků.
Publikováno: 15 listopadu, 2021
více...
Novinky v Průhonické botanické zahradě
Průhonická botanická zahrada leží na východním okraji Průhonického parku. Specializuje se především na konzervaci prastarých a historických odrůd kosatců, pivoněk, denivek a růží.
Publikováno: 15 listopadu, 2021
více...
Vpády botaniků do online výuky škol během covidu
Projekt Vědci do výuky vznikl podobně spontánně jako loni na jaře Věda na doma (https://www.avcr.cz/cs/pro-verejnost/veda-na-doma). Účastnil se ho i Botanický ústav AV ČR.
Publikováno: 15 listopadu, 2021
více...
Vědci dětem / Tohle jsme my…
Pojďme se podívat, jak se vědci z Botanického ústavu věnovali dětem na letním příměstském táboře v Průhonickém parku.
Publikováno: 15 listopadu, 2021
více...

Vyšlo první číslo v roce 2021
S příchodem jara rozkvétají květy na podzámeckém Alpinu a uvolňují pyl. Víte ale vlastně, jak taková pylová zrna vypadají pod mikroskopem? A napadlo vás, jak se pyl chytá? Připojujeme pylový kalendář, který možná ocení nejeden alergik trpící sezonní polinózou.
Publikováno: 15 dubna, 2021
více...
Pylové depozice na Šumavě a v Krkonoších za posledních 21 let
Vědci zabývající se pylem (palynologové) potřebují vědět, v jakém vztahu je spad (depozice) pylu a okolní vegetace. Proporcionální druhové složení pylu známe z hlubokých rašelinných profilů, ale chceme vědět, jaké bylo proporcionální složení vegetace v minulých 10 tisíci letech v...
Publikováno: 15 dubna, 2021
více...
Po těžbě rašeliny (s)poušť?
V českých zemích těžba rašeliny, ale i jiné hospodářské využívání rašelinišť začalo ve srovnání se západní Evropou poměrně pozdě díky relativnímu dostatku lesů a palivového dřeva. Omezení používání dřeva jako paliva a jeho nahrazení uhlím a rašelinou nařizoval pro české...
Publikováno: 15 dubna, 2021
více...
Úhory – zapomenuté ostrůvky biodiverzity
V oblasti Znojemska na jižní Moravě se vyskytují velké rozlohy polí, které výrazně kontrastují s mozaikou drobných políček na rakouské straně hranice. Po obou stranách této hranice se nachází náš nejmenší národní park – Podyjí. Před dvaceti lety začala postupně...
Publikováno: 15 dubna, 2021
více...
Hledání odpovědi na otázku rozmanitosti rostlin na suchozemských ostrovech
V ekologii je velmi populární zkoumat ostrovy a souostroví v rozmanitých souvislostech. Lze na nich testovat ekologické hypotézy týkající se šíření rostlin na velké vzdálenosti, lokální adaptace a vzniku druhů, a to vše ve vztahu ke vzdálenosti ostrova od pevniny,...
Publikováno: 15 dubna, 2021
více...
Paměti rostlin
Také vás při pohledu na starobylý strom napadne, co všechno asi pamatuje? Věřím, že si tuto otázku kladete spíše v kontextu toho, co asi všechno ten úchvatný strom zažil a přežil, než že by si skutečně něco pamatoval. Stromy, a...
Publikováno: 15 dubna, 2021
více...
Vyšlo druhé číslo časopisu Botanika v roce 2020
V druhém čísle časopisu Botanika v roce 2020 se např. dozvíte, proč jsou staré bučiny tak bohaté na lišejníky, dále zajímavosti o kopřivách, za invazními dírkovci se ponoříme pod hladinu moře a po turistických cestách vystoupáme za proměnami krkonošské tundry....
Publikováno: 15 dubna, 2021
více...
Buk – nejvýznamnější česká dřevina pro lišejníky?
Buk lesní (Fagus sylvatica) a lesy jím tvořené není třeba příliš představovat. Buk je rozšířen ve většině Evropy a je často dominantní součástí lesních porostů. Odhady jeho přirozeného zastoupení (tzn. bez změny druhové skladby způsobené člověkem) v českých lesích se...
Publikováno: 15 dubna, 2021
více...
Všudypřítomná, užitečná, ale i zatracovaná kopřiva dvoudomá
Není snad nikdo, kdo by neznal kopřivu a její žahavé chlupy. Proč kopřiva pálí a způsobuje bolestivé puchýře? Při odlomení špičky dutého chlupu dochází ke vzniku ostrého hrotu podobného injekční jehle a vylití pálivé tekutiny. V poslední době se usuzuje,...
Publikováno: 15 dubna, 2021
více...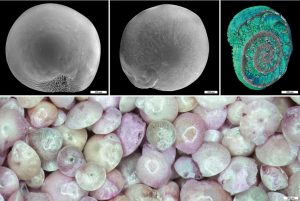
Mrtvé písky Středozemního moře aneb bioeroze schránek invazních dírkovců
Zvídavý čtenář asi zpozorní – proč by měly být písky mrtvé? Nebo ještě zvídavěji – o jaké písky se vlastně jedná? Nebo úplně od začátku: co je to písek? Či zcela logicky a jaksi z druhé strany – existují snad...
Publikováno: 15 dubna, 2021
více...
Kdy trápí pyl alergiky?
Alergické nebo přecitlivělé reakce jsou výsledkem nezdravé imunitní odpovědi k cizím substancím – alergenům, které jsou pro většinu populace neškodné. Původcem pylové alergie (polinózy) je vdechnutý pyl a příčinou jsou alergeny v pylových zrnech obsažené. Tento typ alergie v Evropě...
Publikováno: 15 dubna, 2021
více...
Rostlina roku: vstavač kukačka
Česká botanická společnost se rozhodla každoročně vyhlašovat Rostlinu roku, a to právě od letoška. Inspirací byla akce Pták roku, která je již od roku 1992 organizována Českou společností ornitologickou.
Publikováno: 15 dubna, 2021
více...
Botanické zahrady jako součást evropského kulturního dědictví
Botanické zahrady se celosvětově věnují konzervaci genofondu rostlin. Kromě planých druhů se soustředí také na odrůdy okrasných rostlin, které můžeme chápat jako kulturní dědictví.
Publikováno: 15 dubna, 2021
více...
Jsou vysokohorská společenstva v ohrožení?
Vysokohorské oblasti nabízejí díky interakcím mezi drsným klimatem a členitým reliéfem velkou různorodost stanovišť, na něž jsou vázána úzce specializovaná společenstva rostlin i živočichů (často hmyzu). V našem příspěvku se však budeme věnovat výhradně flóře. Vysokohorské rostliny musí být dobře...
Publikováno: 15 dubna, 2021
více...
Fotografický průvodce Průhonickým parkem – Listnaté stromy 1
Fotografie a mapy ve speciálním čísle časopisu Botanika vám pomohou poznat listnaté stromy v první části Průhonického parku. Znali jste třeba takovou davidii listenovou? Strom z obálky časopisu najdete na velkém nádvoří, kousek za vstupem do Průhonického parku.
Publikováno: 6 května, 2020
více...